
J. T. Thorson1, M. Fossheim2, F. J. Mueter3, E. Olsen4, R. R. Lauth1, R. Primicerio5, B. Husson2, J. Marsh3, A. Dolgov6,7,8, and S. G. Zador1
1Alaska Fisheries Science Center, NOAA, Seattle, WA, USA
2Institute of Marine Research, Tromsø, Norway
3College of Fisheries and Ocean Sciences, University of Alaska Fairbanks, Juneau, AK, USA
4Institute of Marine Research, Bergen, Norway
5University of Tromsø, Tromsø, Norway
6Polar Branch of the Federal State Budget Scientific Institution “Russian Federal Research Institute of Fisheries and Oceanography” (PINRO), Murmansk, Russia
7Federal State Educational Institution of Higher Education “Murmansk State Technical University” (FSEI HE MSTU), Murmansk, Russia
8Tomsk State University, Tomsk, Russia
Highlights
- The northern edge of distribution for fishes associated with the Bering Sea shelf (e.g., walleye pollock and Pacific cod) shifted northward from 2010 to 2017/18.
- The spatial distribution of boreal fishes in the Barents Sea in 2017 resembles their distribution in 2012, showing a northward shift relative to their previous distribution in 2004.
- In both Bering and Barents Seas, recent shifts are associated with changes in bottom water temperature and loss of sea ice.
- Density estimates for individual species show that responses vary among species, and boreal species can maintain high densities in southern habitats even during shifts in the northward edge of distribution.
Introduction
Marine populations are expected to remain within their preferred thermal conditions, and therefore to shift their spatial distributions to track changes in ocean temperatures (Pinsky et al. 2013). Many different indicators show changes in Arctic physical conditions, with an increased rate of change from 2005 to present day (Overland et al. 2019). Given these rapid physical changes and expected responses of marine populations to changing thermal conditions, the spatial distribution of Arctic and subarctic fish communities will likely be a sensitive indicator for contemporary and ongoing Arctic climate change.
Here we present changes in the fish communities of the Bering and Barents Seas during 2017/18 relative to baseline conditions 10-15 years earlier. This involves mapping shifts in the spatial distribution of boreal (southern) and Arctic (northern) fish communities in each ecosystem, as well as identifying indicator species associated with each community. The Bering Sea has shown substantial and rapidly warming ocean temperatures and associated decreases in the thickness and extent of sea ice from 1982 to 2019, with pronounced changes occurring since 2014, while the sea surface temperature trends in the Barents Sea are varied both spatially and seasonally (see essay Sea Surface Temperature). Fish populations in both the Bering and Barents Seas are expected to expand into new areas in a warming climate if certain pre-conditions are met, including suitable bottom topography, water temperatures, salinities and distance to spawning grounds (Hollowed et al. 2013). The patterns and rates of documented shifts are species-specific and tend to follow temperature gradients (Pinsky et al. 2013), although observed shifts have trailed behind and are sometimes poorly correlated with the actual rates of climate shifts (Alabia et al. 2018).
Bering Sea community shifts
On the shallow Bering Sea shelf, the summer distribution of fishes and invertebrates living near the sea floor (i.e., demersal fishes) is tied to the extent of the cold pool (bottom temperatures < 2°C) as determined by the southern extent of sea ice during the preceding winter (Mueter and Litzow 2008). Until recently, there was a general expectation that sea ice would continue to persist throughout the winter over the shallow northern Bering Sea and Chukchi Sea shelf, limiting the northward expansion of boreal species (Stabeno et al. 2012). However, latent heat from the warm conditions of summer 2016, combined with less sea-ice formation in the 2017 winter, resulted in an extensive but narrow cold pool on the eastern Bering Sea shelf during summer 2017 (see essay Recent Warming in the Bering Sea). This created a wide corridor within the shallow continental shelf for species to move northward, as observed with dramatic increases of Pacific cod, Gadus macrocephalus, walleye pollock, G. chalcogrammus and several flatfish species into the northern Bering Sea (Stevenson and Lauth 2019). A delayed freeze-up in the Chukchi Sea and unusual southerly winds resulted in near-complete lack of sea ice in the northern Bering Sea the following winter, in 2018. Maximum sea-ice extent subsequently reached the lowest levels on record in 2019 (see essay Recent Warming in the Bering Sea).
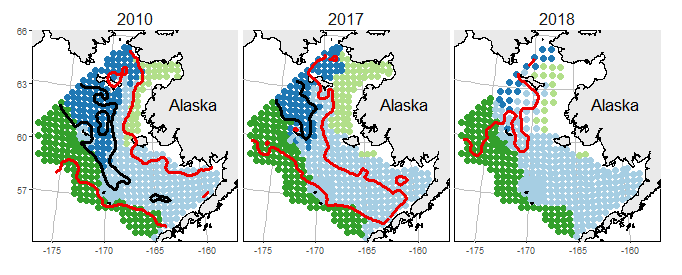
We identify shifts in distribution for four assemblages in the eastern and northern Bering Sea (Fig. 1), and also identify indicator species that are associated with each assemblage (Table 1). Boreal groundfish were observed again in the northern Bering Sea during the 2018 summer (Duffy-Anderson et al. 2019; Stabeno et al. 2019). Although shifts in these large, abundant species are more apparent, Arctic taxa tend to be more sensitive to habitat changes. Arctic cod (Boregadus saida) in particular can serve as a sentinel species that responds quickly to changes in temperature and ice extent (Alabia et al. 2018; Marsh and Mueter 2019). Changes in individual species result in community-level changes (Fig. 1) that reflect a transition from a ‘Northern Shelf’ (Arctic) community dominated by relatively few, smaller and less abundant species with Arctic affinities to a community dominated by larger boreal species. In particular, the Arctic and northern shelf community is represented by butterfly sculpin, Hemilepidotus papilio, and Bering flounder, Hippoglossoides robustus (see Table 1 for full list), and this community retreated northward from 2010 to 2018. This Arctic community was then replaced by the southern shelf community represented by northern rock sole, Lepidopsetta polyxystra, and sturgeon poacher, Podothecus accipenserinus (i.e., comparing the spatial distribution of light and dark blue in Fig. 1).
| Community | Species | |
|---|---|---|
| Scientific Name | Common Name | |
| Boreal: outer shelf / slope | Atheresthes evermanni | Kamchatka flounder |
| Atheresthes stomias | arrowtooth flounder | |
| Lycodes brevipes | shortfin eelpout | |
| Hippoglossoides elassodon | flathead sole | |
| Hemitripterus bolini | bigmouth sculpin | |
| Icelus spiniger | thorny sculpin | |
| Chionoecetes bairdi | tanner crab | |
| Bathyraja interrupta | sandpaper skate | |
| Glyptocephalus zachirus | rex sole | |
| Reinhardtius hippoglossoides | Greenland halibut/turbot | |
| Leptagonus frenatus | sawback poacher | |
| Boreal: southern shelf | Lepidopsetta polyxystra | Northern rock sole |
| Podothecus accipenserinus | sturgeon poacher | |
| Paralithodes camtschaticus | red king crab | |
| Norton Sound | Eleginus gracilis | saffron cod |
| Myoxocephalus jaok | plain sculpin | |
| Platichthys stellatus | starry flounder | |
| Limanda proboscidea | longhead dab | |
| Gymnocanthus pistilliger | threaded sculpin | |
| Telmessus cheiragonus | helmet crab | |
| Lumpenus fabricii | slender eelblenny | |
| Arctic: northern shelf | Hemilepidotus papilio | butterfly sculpin |
| Liparis gibbus | variegated snailfish | |
| Limanda sakhalinensis | Sakhalin sole | |
| Hippoglossoides robustus | Bering flounder | |
| Boreogadus saida | Arctic cod | |
| Lycodes raridens | marbled eelpout | |
| Myoxocephalus verrucosus | warty eelpout | |
Barents Sea community shifts
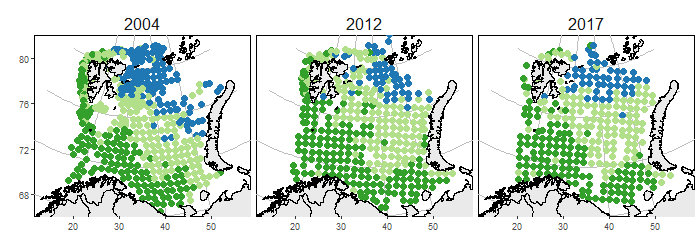
In the Barents Sea, the warming ocean temperatures and retraction of the sea ice was concurrent with a northeastward displacement of Arctic fish (Fig. 2). This region includes three communities: Arctic (represented by Arctic cod, Boreogadus saida and bigeye sculpin, Triglops nybelini), boreal (represented by haddock, Melanogrammus aeglefinus and saithe, Pollachius virens) and a transition community (see Table 2 for full list of species associated with each community). The decreasing extent of Arctic water masses was associated with a northward shift in the transition zone separating Arctic and boreal fish communities from 2004 to 2012 (Fossheim et al. 2015a,b). During these years, the southern (boreal) fish community gradually expanded northward while the northern (Arctic) community retracted (Fossheim et al. 2015a,b). In 2017 the Arctic fish communities continued to have a displaced spatial distribution similar to 2012; there was neither a recovery to previous distributions (like 2004), nor a further displacement (beyond 2012). The boreal community includes large-bodied generalist feeders. These species took advantage of favorable conditions in the Barents Sea, both by increasing in abundance (demography) and by expanding their distribution (behavior). By contrast, many Arctic species are small-sized bottom-associated specialists and their spatial distribution has retracted to smaller areas in the northeast. We hypothesize that new areas for boreal species opened as Arctic waters warmed and primary and secondary production increased (see essay Arctic Ocean Primary Productivity). These changes likely favored the boreal generalists competitively over the arctic specialists. In some cases, the larger predatory species (i.e., boreal fish) might also exert feeding pressure on the smaller benthivore species (i.e., Arctic fish).
| Community | Species | |
|---|---|---|
| Scientific Name | Common Name | |
| Boreal | Melanogrammus aeglefinus | Haddock |
| Micromesistius poutassou | blue whiting | |
| Sebastes mentella | deepwater redfish | |
| Amblyraja radiata | thorny skate | |
| Sebastes norvegicus | golden redfish | |
| Trisopterus esmarkii | Norway pout | |
| Anarhichas minor | spotted wolffish | |
| Pollachius virens | saithe | |
| Mixed | Leptoclinus maculatus | daubed shanny |
| Lumpenus lampretaeformis | snakeblenny | |
| Triglops murrayi | moustache sculpin | |
| Arctic | Triglops nybelini | bigeye sculpin |
| Boreogadus saida | Arctic cod | |
| Liparidae spp. | snailfish spp. | |
| Reinhardtius hippoglossoides | Greenland halibut/turbot | |
| Icelus spp. | sculpin spp. | |
Single-species analyses
We next illustrate these same patterns for individual, large-bodied species in each region. For walleye pollock and Pacific cod in the eastern Bering Sea, northward shifts in distribution are clearly apparent between bottom-trawl surveys occurring first in 2010 and repeated in 2017 and 2018 (Fig. 3a-f). Hotspots of increased density for Pacific cod are apparent south of St. Lawrence Island (63° N -170° W) and for walleye pollock along the limits of the Russian exclusive economic zone near the Bering Strait (64° N -172° W) in 2017/18. The locations of these hotspots along the northern portion of the Bering Sea have contributed to a rapid northward shift in the population centroid for these and other commercially important species (e.g., northern rock sole, Lepidopsetta polyxystra, see Stevenson and Lauth (2019)). Distribution shifts in the Barents Sea are also apparent by comparing densities of Atlantic cod and haddock in the summer bottom-trawl survey from 2003 with surveys in 2017/18 (Fig. 3g-l). For example, Atlantic cod show increased densities in the central Barents Sea (75° N 35° E) while haddock show increased densities near Bear Island (75° N 19° E). These species-specific results show that the areas with the greatest changes vary from species to species. These results also show that northward shifts can occur even while population densities for boreal species remain high within the southern portion of their range, e.g., elevated estimates of walleye pollock density near Unimak Island (65° N -165° W) in 2010 and 2018 (Fig. 3a,c).

Consequences for Arctic ecosystems
In the Bering Sea, the expansion of nearshore (‘Norton Sound’) and southern shelf species into the northern Bering Sea is likely to have profound effects on the benthic ecology of the northern Bering Sea, due to an increase in demersal fish biomass by several orders of magnitude (Stevenson and Lauth 2019). The observed high densities of fish in Bering Strait suggest that boreal species may shift their summer feeding migrations into the Chukchi Sea, which has also shown large decreases in summer sea-ice extent (see essay Sea Ice). Movement northward into the Chukchi Sea highlights the need for additional fisheries surveys north of Bering Strait, ongoing monitoring of benthic ecosystem components in the northern Bering and Chukchi seas, as well as synthesis of data from the western portion of the Bering Sea within Russian waters. In the Barents Sea, fish communities shifted rapidly from 2004 to 2012, and then remained relatively stable from 2012 to 2017. Increasing water temperatures opened new feeding habitats in the Barents Sea for Atlantic cod and haddock. So far, these species have been benefiting from the effects of climate change on the Barents Sea. These recent bottom-trawl surveys suggest that the spatial distributions of fish communities in the Barents Sea are stable for the moment at a new state, associated with warmer waters, less sea ice, and northward displacement of boreal species. However, different processes drive density changes in leading (northern) and trailing (southern) edges of species distribution (Pinsky et al. 2020), and contemporary warming could still result in future (lagged) changes. We encourage further comparison between climate-driven distribution shifts for these Atlantic and Pacific gateways to the Arctic. In particular, we note the importance of separating mechanisms occurring on the leading (northward) edges where populations are colonizing new habitats, relative to trailing (southward) edges where populations are likely to be extirpated from existing habitat given future warming.
Acknowledgments
We thank the many scientists who have contributed to these long-term surveys of the eastern and northern Bering Sea, and the Barents Sea. We also thank L. Britt, L. Barnett, E. Yasumiishi, and three anonymous reviewers for comments on an earlier draft.
References
Alabia, I. D., J. G. Molinos, S. -I. Saitoh, T. Hirawake, T. Hirata, and F. J. Mueter, 2018: Distribution shifts of marine taxa in the Pacific Arctic under contemporary climate changes. Divers. Distrib., 24, 1583-1597, https://doi.org/10.1111/ddi.12788.
Duffy-Anderson, J. T., and Coauthors, 2019: Responses of the northern Bering Sea and southeastern Bering Sea pelagic ecosystems following record-breaking low winter sea ice. Geophys. Res. Lett., 46(16), 9833-9842, https://doi.org/10.1029/2019GL083396.
Dufrêne, M., and P. Legendre, 1997: Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr., 67, 345-366, https://doi.org/10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2.
Fossheim, M., R. Primicerio, E. Johannesen, R. B. Ingvaldsen, M. M. Aschan, and A.V. Dolgov, 2015a: Climate change is pushing boreal fish northwards to the Arctic: The case of the Barents Sea. Arctic Report Card 2015, M. O. Jeffries, J. Richter-Menge, and J. E. Overland, Eds., https://www.arctic.noaa.gov/Report-Card.
Fossheim, M., R. Primicerio, E. Johannesen, R. B. Ingvaldsen, M. M. Aschan, and A.V. Dolgov, 2015b: Recent warming leads to a rapid borealization of fish communities in the Arctic. Nat. Climate Change, 5, 673-677, https://doi.org/10.1038/nclimate2647.
Hollowed, A. B., and Coauthors, 2013: Projected impacts of climate change on marine fish and fisheries. ICES J. Mar. Sci., 70, 1023-1037, https://doi.org/10.1093/icesjms/fst081.
Marsh, J. M., and F. J. Mueter, 2019: Influences of temperature, predators, and competitors on polar cod (Boreogadus saida) at the southern margin of their distribution. Polar Biol., https://doi.org/10.1007/s00300-019-02575-4.
Mueter, F.J., and M. A. Litzow, 2008: Sea ice retreat alters the biogeography of the Bering Sea continental shelf. Ecol. Appl., 18, 309-320, https://doi.org/10.1890/07-0564.1.
Overland, J. E., M. Wang, and J. E. Box, 2019: An integrated index of recent pan-Arctic climate change. Environ. Res. Lett., 14, 035006, https://doi.org/10.1088/1748-9326/aaf665.
Pinsky, M. L., B. Worm, M. J. Fogarty, J. L Sarmiento, and S. A. Levin, 2013: Marine taxa track local climate velocities. Science, 341, 1239-1242.
Pinsky, M. L., R. L. Selden, and Z. J Kitchel, 2020 (in press): Climate-driven shifts in marine species ranges: Scaling from organisms to communities. Annu. Rev. Mar. Sci., 12, https://doi.org/10.1146/annurev-marine-010419-010916.
Stabeno, P. J., E. V. Farley Jr., N. B. Kachel, S. Moore, C. W. Mordy, J. M. Napp, J. E. Overland, A. I. Pinchuk, and M. F. Sigler, 2012: A comparison of the physics of the northern and southern shelves of the eastern Bering Sea and some implications for the ecosystem. Deep-Sea Res. Pt. II, 65-70, 14-30, https://doi.org/10.1016/j.dsr2.2012.02.019.
Stabeno, P. J., S. W. Bell, N. A. Bond, D. G. Kimmel, C. W. Mordy, and M. E. Sullivan, 2019: Distributed biological observatory region 1: Physics, chemistry and plankton in the northern Bering Sea. Deep Sea Res. Pt. II, 162, 8-21, https://doi.org/10.1016/j.dsr2.2018.11.006.
Stevenson, D. E., and R. R. Lauth, 2019: Bottom trawl surveys in the northern Bering Sea indicate recent shifts in the distribution of marine species. Polar Biol., 42, 407-421, https://doi.org/10.1007/s00300-018-2431-1.
Thorson, J. T., 2019: Guidance for decisions using the Vector Autoregressive Spatio-Temporal (VAST) package in stock, ecosystem, habitat and climate assessments. Fish. Res., 210, 143-161, https://doi.org/10.1016/j.fishres.2018.10.013.
November 19, 2019